
Cell size and organ size
Size is a fundamental characteristic of every organism; animals from mouse to elephant and plants from Wolfia, the smallest flowering plant, to the towering giant sequoia trees must establish and maintain proper organ size and shape. Peter Lawrence has remarked that size “is the material that evolution largely works on” (Vogel, 2013). How an organ senses its size and determines when to stop growing is one of the biggest remaining mysteries in developmental biology, as has been acknowledged recently in Science (Vogel, 2013). Developmental processes regulating size and shape are remarkably reliable in producing nearly invariant organs within a species. For example, the two arms of a person match in length with an accuracy of 0.2% (Wolpert, 2010), and Arabidopsis floral organs are strikingly uniform in size and shape (Mizukami, 2001). The size of an organ is determined by the number and size of its cells. Confoundingly, the behavior of these cells is often variable and unpredictable. Equivalent neighboring plant cells grow at markedly different rates in several developing tissues (Armour et al., 2015; Elsner et al., 2012; Kierzkowski et al., 2012; Tauriello et al., 2015; Uyttewaal et al., 2012); although at later stages of development, growth may become more uniform (Zhang et al., 2011). Similarly, neighboring cells have different constriction rates during Drosophila gastrulation (Martin et al., 2009). The research in my laboratory falls at the intersection of cell biology and developmental biology, focusing on two interrelated fundamental questions: (1) how are variable cell sizes produced during development and (2) how do reproducible organ sizes and shapes emerge from the variable growth of their cells? The theme emerging from my lab’s research is the importance of cellular variability in plant development. This principle is surprising because biologists generally think of development as being a highly regulated and reproducible process, where variability must be suppressed. Instead we are finding that the plant utilizes this variability to produce regularity.
Computational Morphodynamics Approach—live imaging, image analysis, and computational modeling

To answer these complex questions about cell size and organ size, we take an interdisciplinary collaborative approach, working with physicists, computer scientists, and applied mathematicians who have a particular interest in plant biology. With this team we reach conclusions that could not be reached from any one of these perspectives alone. These collaborations are intensive—we meet every two to three weeks through videoconferencing and exchange many e-mails in between. Experimenters in my lab are intimately involved in designing the analysis and models of our collaborators, and our collaborators are intimately involved in guiding our next experiments. In my lab, we use molecular genetic approaches in Arabidopsis thaliana to identify new genes and their functions in variable cell size and regular organ size and elucidate how these genes relate to one another in a genetic network. To analyze cell division, growth, gene expression, and their relationship to organ growth, we image the same living organ every 8 to 12 hours on a confocal microscope, in a technique called live imaging. This approach allows us to observe the development of the organ at cellular resolution (Hervieux et al., 2016). We then use sophisticated image analysis computer programs developed by our collaborators to detect each cell, measure its growth rate, growth direction, and cell division, and quantify fluorescent protein expression over time (Roeder et al., 2012a). In particular we use the MorphoGraphX package developed by our collaborator Richard Smith (Barbier de Reuille et al., 2015). Collaborators Gerardo Tauriello, Petros Koumoutsakos and I developed the “parent labels” function of this software package that allows us to track a cell and each of its daughters over time, which forms the basis for all of our analysis of growth and division (Tauriello et al., 2015). Simultaneous with the data generation and analysis, our collaborators Arezki Boudaoud, Henrik Jönsson, and James Locke build computational models reflecting our hypotheses about the underlying mechanisms. Often the model does not match the data initially, leading us to perform additional experiments, modify our hypotheses, and revise our model until the data and the model match. This combined interdisciplinary interaction between experiments, particularly using live imaging, image analysis, and computational modeling constitutes a computational morphodynamics approach (Chickarmane et al., 2010; Roeder et al., 2011).
Novel Arabidopsis sepal model system for cell and organ size research

Members of my laboratory and I have developed the Arabidopsis thaliana sepal, the outermost green leaf-like floral organ, as a model system for investigating cellular variability and organ regularity. The sepal is the outermost, green, leaf-like floral organ, which encloses and protects the developing reproductive organs. The sepal is advantageous because it is accessible at all developmental stages for imaging and manipulation. The sepal is relatively small, so the entire development of the organ can be imaged on a confocal microscope, allowing live imaging of the same sepal throughout its development.
I. Variable cell size: The pattern of giant cells and small cells in the Arabidopsis sepal
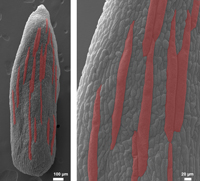
The Arabidopsis sepal is an ideal system for studying developmental regulation producing cell size variability because the cells in the outer (abaxial) epidermis are diverse in size, ranging from giant cells stretching to an average of 360 µm in length to the smallest cells reaching only about 10 µm in length (Roeder et al., 2010). Giant cells form scattered among smaller cells. One function of giant cells is to control the curvature of the organ, which is essential for the sepal’s protective function. Sepal curvature is altered in mutants with too few or too many giant cells, suggesting that these cells have a role in morphogenesis. In the sepal, cell size is correlated with the type of cell cycle: division or endoreduplication. Endoreduplication is a specialized cell cycle in which a cell replicates its DNA, but does not divide, thus becoming enlarged and polyploid. Giant cells undergo three rounds or four rounds of endoreduplication, to become endopolyploid 16C or 32C cells (Roeder et al., 2010). Small cells undergo only mitotic cell cycles and remain generally 2C or 4C. There are 8C cells of intermediate size that have endoreduplicated twice. Two molecular markers show that giant cells and small cells have distinct patterns of gene expression as well as distinct cell sizes and DNA contents. Endoreduplicated tissues have agricultural importance; highly endoreduplicated tissues, such as cotyledons in peas and beans and endosperm in grains, such as wheat, corn, and rice, are used to store nutrition. However, these structures all develop deep within the seed coat and are inaccessible for observation and manipulation. The Arabidopsis sepal, in which endoreduplication occurs in the outer surface cells that are visible in confocal microscopy, is a good model system for examining endoreduplication during development.
We have found that the formation of giant cells and their endoreduplication is regulated by developmental control genes in the epidermal specification pathway, including Arabidopsis thaliana MERISTEM LAYER1 (ATML1) (Roeder et al., 2012b). We have further found an unexpected, yet specific, role of the endomembrane trafficking protein SEC24A in endoreduplication and cell size patterning in the Arabidopsis sepal (Qu et al., 2014). Fluctuations in the transcription factor Arabidopsis thaliana MERISTEM LAYER1 (ATML1) pattern giant cells and small cells in the Arabidopsis sepal epidermis (Meyer et al., 2017). We further show that the G2 stage of the cell cycle provides a permissive window when cells can be specified as giant due to high levels of ATML1. If a cell reaches a high concentration of ATML1 during the G2 stage, it is likely to endoreduplicate and become a giant cell. If the ATML1 concentration remains below the threshold, the cell is likely to divide. Thus, we identify a novel fluctuation-dependent patterning mechanism; our results suggest that random variation in the expression level of a key transcription factor can be an important component of producing highly tuned developmental patterns (Meyer et al., 2017). This result points to the importance of variability in plant development.
II. Reproducible organ size: Production of strikingly reproducible organ sizes and shapes from the variable growth of their cells
Arabidopsis floral organs, particularly sepals, allow robustness in organ size and shape to be assessed within a single plant. Each flower has four sepals of the same size, and individual plants can produce more than 100 flowers, allowing a statistical assessment of organ size uniformity within an individual organism, which generally cannot be done in animals. The consistent size and shape of sepals is required to enclose and protect the developing reproductive organs, maintaining an effective barrier against the external environment. In addition, the size of floral organs is relatively insensitive to environmental effects, allowing us to focus on intrinsic mechanisms of size control. Uniformity in size, shape, and time of ripening are a critical for efficient harvest, transportation and marketing of many agricultural products including fruits and vegetables (Hotegni et al., 2014).
We have found that sepal cell growth rate is highly variable (Tauriello et al., 2015). Underlying this variable cell growth there is a general pattern in which cells mature, slow their growth, and stop dividing progressively from the tip to the base (basipetal) of the sepal (Hervieux et al., 2016). We further found that the formation of characteristic nano-ridge structures in the cuticle coincides with the wave of maturation (Hong et al., 2017). We have found that spatiotemporal averaging of cell growth variability promotes organ regularity and that the ftsh4 mutant with reduced cell variability has irregular size and shape sepals (Hong et al., 2016). Spatiotemporal averaging works for example if initially cell A is growing fast and its neighbor cell B is growing slowly, then later cell A is growing slowly and cell B is growing fast, their growth averages out to a uniform moderate growth rate. Thus, we are working on elucidating the complex relationship between cell growth and organ growth, which is not well understood in plants or animals.
III. Polyploidy Integration and Innovation Institute (PI3)
Extending from our work on giant cells and endoreduplication, we are now working on understanding the fundamental roles of genome doubling on the cell, the transcriptome, the proteome, how this influences the performance of the tissue, the organism, the population and the landscape. Our role is focusing on cell to tissue to organism in Arabidopsis. Our goal is to compare across organisms including Drosophila, yeast, chlamydomonas, and duckweed with our team in the NSF-funnded BII Polyploidy Integration and Innovation Institute (PI3). More information can be found at our PI3 website.
IV. Engineered Living Materials (ELM)
We are part of the Engineered Living Materials Institute (ELMI) at Cornell. We are applying our knowledge of plant morphogenesis to investigating how to grow sustainable materials from living organisms. We currently have a projects diatoms and cell wall mechanics.
References:
Armour, W.J., Barton, D.A., Law, A., and Overall, R.L. (2015). Differential growth in periclinal and anticlinal walls during lobe formation in Arabidopsis cotyledon pavement cells. Plant Cell 27, 2484–2500.
Barbier de Reuille, P., Routier-Kierzkowska, A.L., Kierzkowski, D., Bassel, G.W., Schüpbach, T., Tauriello, G., Bajpai, N., Strauss, S., Weber, A., Kiss, A., et al. (2015). MorphoGraphX: A platform for quantifying morphogenesis in 4D. eLife 4, e05864.
Chickarmane, V., Roeder, A.H.K., Tarr, P.T., Cunha, A., Tobin, C., and Meyerowitz, E.M. (2010). Computational morphodynamics: a modeling framework to understand plant growth. Annual Review of Plant Biology 61, 65–87.
Elsner, J., Michalski, M., and Kwiatkowska, D. (2012). Spatiotemporal variation of leaf epidermal cell growth: a quantitative analysis of Arabidopsis thaliana wild-type and triple cyclinD3 mutant plants. Ann Bot 109, 897–910.
Hervieux, N., Dumond, M., Sapala, A., Routier-Kierzkowska, A.-L., Kierzkowski, D., Roeder, A.H.K., Smith, R.S., Boudaoud, A., and Hamant, O. (2016). A mechanical feedback restricts sepal growth and shape in Arabidopsis. Curr Biol 26, 1019–1028.
Hong, L., Dumond, M., Tsugawa, S., Sapala, A., Routier-Kierzkowska, A.-L., Zhou, Y., Chen, C., Kiss, A., Zhu, M., Hamant, O., et al. (2016). Variable Cell Growth Yields Reproducible Organ Development through Spatiotemporal Averaging. Dev Cell 38, 15–32.
Hong, L., Brown, J., Segerson, N.A., Rose, J.K.C., and Roeder, A.H.K. (2017). CUTIN SYNTHASE 2 Maintains Progressively Developing Cuticular Ridges in Arabidopsis Sepals. Mol Plant 10, 560–574.
Hotegni, V., Lommen, W., and Agbossou, E.K. (2014). Heterogeneity in pineapple fruit quality results from plant heterogeneity at flower induction. Front Plant Sci 5, 670.
Kierzkowski, D., Nakayama, N., Routier-Kierzkowska, A.-L., Weber, A., Bayer, E., Schorderet, M., Reinhardt, D., Kuhlemeier, C., and Smith, R.S. (2012). Elastic domains regulate growth and organogenesis in the plant shoot apical meristem. Science 335, 1096–1099.
Martin, A.C., Kaschube, M., and Wieschaus, E.F. (2009). Pulsed contractions of an actin-myosin network drive apical constriction. Nature 457, 495–499.
Meyer, H.M., Teles, J., Formosa-Jordan, P., Refahi, Y., San-Bento, R., Ingram, G., Jönsson, H., Locke, J.C.W., and Roeder, A.H.K. (2017). Fluctuations of the transcription factor ATML1 generate the pattern of giant cells in the Arabidopsis sepal. eLife 6, 635–641.
Mizukami, Y. (2001). A matter of size: developmental control of organ size in plants. Curr Opin in Plant Biol 4, 533–539.
Qu, X., Chatty, P.R., and Roeder, A.H.K. (2014). Endomembrane trafficking protein SEC24A regulates cell size patterning in Arabidopsis. Plant Phys 166, 1877–1890.
Roeder, A.H.K., Chickarmane, V., Cunha, A., Obara, B., Manjunath, B.S., and Meyerowitz, E.M. (2010). Variability in the control of cell division underlies sepal epidermal patterning in Arabidopsis thaliana. PLoS Biol 8, e1000367.
Roeder, A.H.K., Cunha, A., Burl, M.C., and Meyerowitz, E.M. (2012a). A computational image analysis glossary for biologists. Development 139, 3071–3080.
Roeder, A.H.K., Cunha, A., Ohno, C.K., and Meyerowitz, E.M. (2012b). Cell cycle regulates cell type in the Arabidopsis sepal. Development 139, 4416–4427.
Roeder, A.H.K., Tarr, P.T., Tobin, C., Zhang, X., Chickarmane, V., Cunha, A., and Meyerowitz, E.M. (2011). Computational morphodynamics of plants: integrating development over space and time. Nat Rev Mol Cell Biol 12, 265–273.
Tauriello, G., Meyer, H.M., Smith, R.S., Koumoutsakos, P., and Roeder, A.H.K. (2015). Variability and constancy in cellular growth of Arabidopsis sepals. Plant Phys 169, 2342–2358.
Uyttewaal, M., Burian, A., Alim, K., Landrein, B., Borowska-Wykręt, D., Dedieu, A., Peaucelle, A., Ludynia, M., Traas, J., Boudaoud, A., et al. (2012). Mechanical stress acts via katanin to amplify differences in growth rate between adjacent cells in Arabidopsis. Cell 149, 439–451.
Vogel, G. (2013). Mysteries of development: How do organs know when they have reached the right size? Science 340, 1156–1157.
Wolpert, L. (2010). Arms and the man: The problem of symmetric growth. PLoS Biol 8, e1000477.
Zhang, C., Halsey, L.E., and Szymanski, D.B. (2011). The development and geometry of shape change in Arabidopsis thaliana cotyledon pavement cells. BMC Plant Biol 11, 27.